Morphology, anatomy, reproduction and interrelationship of Bennettitales: Cycadioidea, Williamsonia
Bennettitales:-















> This group of fossil plants flourished well during the Triassic to Lower Cretaceous periods of Mesozoic era. As the Carboniferous period is called the “Ages of Ferns “, the Mesozoic era is called the ‘Ages of Cycads’. It is due to the fact that Cycadeoideales co-existed with Cycadales during Mesozoic era from Jurassic up to Cretaceous period, and hence this period is called Age of Cycads.
> Bennettitales are found either in the form of compressions or petrifactions. Due to Cycad-like form of their fronds and the presence of short stems covered with an armour of presistent leaf bases Bennettitales (Cycadeoideales) have been treated under Cycadophyta by some workers. However, the two groups are quite distinct from each other and maintain their independent identity.
General Characters:-
1. These extinct Mesozoic plants were present were present on the earth from Triassic to Cretaceous.
2. Bennettitales were so abundant during Mesozoic era that this period is known as ‘Age of Cycads’.
3. The members of this group are found either as compressions or petrifactions.
4. The stems were stout or slender and had a wide pith.
5. The stem grew very slowly and had manoxylic wood.
6. Resembling living Cycads, the Bennettitalean leaves were mostly pinnately compound, and only occasionally simple.
7. Venation was open, and only rarely closed.
8. Syndetocheilic type of stomata were present.
9. The wall of the epidermal cells was sinuous.
10. The reproductive organs were organised in the form of hermaphrodite (e.g. Cycadeoidea) or unisexual (e.g. Wielandiella) “flowers”, which in turn were protected by many bracts.
11. The ‘flowers’ developed in the axil of leaves.
12. Male reproductive organs were borne in a whorl. They were free or fused, entire or pinnately compound.
13. Microsporangia were present abaxially in the form of synangia.
14. Microsporophyll’s sometimes surrounded megasporophylls forming hermaphrodite “flowers”.
15. Ovules were numerous and stalked and borne on a conical, cylindrical or dome-shaped receptacle.
16. Many inter-seminal bracts were present on the ovule containing receptacle.
17. The scales or bracts were united at end to form shield through which micropyle protrudes.
18. Seeds were dicotyledonous.
Interrelationships:-
Resemblances of Bennettitales with Ferns:-
• Bennettitalean plants had multicellular ramenta on their entire body, a characteristic also seen in
ferns.
• Some other features suggesting the filicean affinity of Bennettitales include the presence of:
(i) Direct leaf traces,
(ii) Scalariform tracheids, and
(iii) Large pith.
Resemblances of Bennettitales and Cycads:-
• Bennettitales resemble cycads in the:
(i) structure of their fronds,
(ii) presence of short stems covered with an armour of persistent leaf bases,
(iii) presence of barrel-shaped trunk,
(iv) presence of very thick cortex, relatively thin wood and large pith in the stem,
(v) manoxylic wood,
(vi) monocolpate pollen grains,
(vii) orthotropous ovules, and
(viii) dicotyledonous embryo.
Resemblances between Bennettitales with Pteridospermales:-
• Characters common in both Bennettitales and Pteridospermales include:
(i) Presence of ramenta] hairs,
(ii) Syndetocheilic stomata,
(iii) Direct leaf traces,
(iv) Similar anatomical details,
(v) Leafy microsporophyll’s, and
(vi) Presence of cupule.
• The so-called bisporangiate ‘flower’ of Bennettitales could be compared with the bisporangiate fronds of Ptendospermales.
• Scientists are of the opinion that there exist two lines of evolution from Pteridospermales. Of these, one line gave rise to Bennettitales possessing both uni- and bisporangiate forms, and the other gave rise to mono-sporangiate forms of cycads.
Williamsonia:-
1. Occurrence of Williamsonia:-
> Williamsonia belongs to family Williamsoniaceae of Bennettitales. It has been reported from Upper Triassic period but was more abundant in Jurassic. This was earlier discovered under the name Zamia gigas by Williamson (1870) but has now been named as Williamsonia.
> Professor Birbal Sahni (1932) described W. sewardiana from Rajmahal Hills of Bihar (India). Professor AC. Seward, a well-known palaeobotanist, described W.scotti. Gupta (1943) discovered Williamsonia sahnii from Rajmahal Hills and named if after Professor Birbal Sahni.
> Other reported species from Rajmahal Hills are Williamsonia indica, W. microps and W. santalensis. Bucklandia indica, described from Rajmahal Hills, is now considered to be the stem of Williamsonia sewardiana.
2. External Features of Williamsonia:-
> Williamsonia (Fig. 6.13) resembled Cycas in appearance, and its best known species is W. sewardiana. A reconstruction of this species was published by Sahni (1932). The leaves of W. sewardiana were like that of Ptilophyllum. The plant had an upright, branched and stout stem covered by persistent leaf bases.
> A terminal crown of pinnately compound leaves was present. For the stem genus Bucklandia, Sharma (1991) opined that features of leaf bases such as their shape, size and arrangement pattern are of taxonomic significance.
> He observed that leaves in Williamsoniaceae show syndetocheilic stomata with rachis possessing collateral endarch vascular bundles arranged in a double U-manner. A distinct constriction was present at the base of lateral shoots.

3. Reproduction in Williamsonia:-
> The fructifications of Williamsonia were large and attained a diameter of about 12 cm. They were borne on a peduncle. Many spirally arranged bracts were present around the base of the floral axis. In W. gigas the cones were present among the crown of leaf bases while in W. sewardiana they were present on the short lateral branches. Williamsonia plants were unisexual.
a. Female Flower:-
- The female ‘cones’ of Williamsonia gigas and W. sewardiana have been investigated in detail. Instead of ‘strobili’ or ‘cones’, Sporne (1965) has proposed to use the term “flower” in Williamsonia. The structure of female flower of W. gigas is illustrated in Figs. 6.14, 6.15. The conical receptacle was surrounded by many perianth-like bracts. The ovules were stalked.
- The apex of the receptacle was naked and sterile. The nucellus was surrounded by a single vascularize integument, which was fused with the nucellus. The nucellus had a well-marked beak and a pollen chamber. In young ovules the micropylar canal was long and narrow.
- In mature ovules, the canal widened because of the formation of nucellar plug and disappearance of interlocking cells. In the apical part of endosperm, Sharma (1979) observed 2 or more archegonia.


b. Male Flower:-
- Out of several known male flowers of Bennettitales some have been described to belong to Williamsonia. Male fructifications have never been found in actual connection with the plant, and are sometimes referred to the genus Weltrichia.


- Male flowers consisted of a whorl of microsporophyll’s, which were united to form a more or less cuplike structure. In majority of the investigated species (e.g. Williamsonia whitbiensis) the sporophylls were un-branched but in some species (e.g. W.spectabilis) they were also pinnately branched.

- Sitholey and Bose (1953) discovered Williamsonia santalensis from Upper Gondwana (India), and observed that microsporophyll’s in the species were bifid. One of the branches of microsporophyll was fertile while the other was sterile. The fertile part had finger-like structures called synangia Each synangium had two rows of chambers enclosing microsporangia. Sharma (1977,1983, 1991) has confirmed the synangiate nature.
- The entire male flower attained a length of about 20 cm., while a single microsporophyll was about 10 cm long. The fertile branch of the bifid sporophvll possessed many purse-like capsules, in each of which there were present many monocolpate pollen grains.
Cycadeoidea:-
1. Morphological Features of Cycadeoidea:-
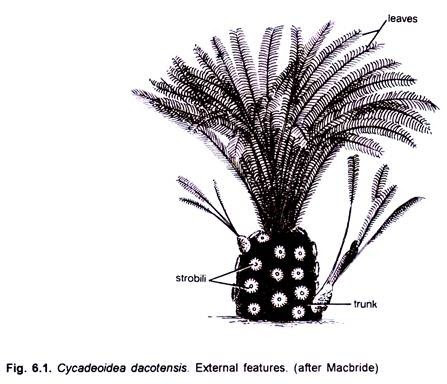
> The Cycadeoid trunks were short, stout, spherical to sub-spherical and un-branched or branched. The trunks and leaves of many of its species show remarkable resemblance with those of living Cycads. Some of the species were short while others (Cycadeoidea jenneyana) attained a height of 3 to 3 .6 metres.
> The trunk generally attained a diameter of about 50 cm, and had many, persistent, rhomboidal leaf bases. A compact crown of Cycad-like, large, pinnately compound leaves was present at the apex. The leaflets had many parallel veins.



2. Anatomy of Cycadeoidea:-
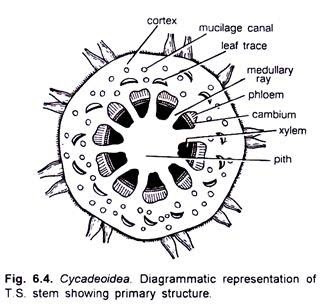
> The stem was roughly circular or oval in outline. It remained covered by heavy armour of leaf bases. The epidermis was not very distinct. The cortex was parenchymatous and possessed many mucilage canals and leaf traces. Many conjoint, collateral, open and endarch vascular bundles constituted the primary vasculature of the stem. A large centrally located pith was present.

> The xylem and the pholem have been studied in detail by Wieland (1906) (Fig. 6.5 A, C) Most of the tracheids were rectangular in shape. They were scalariform. The tracheids of protoxylem were spiral. The secondary xylem and the secondary phloem were traversed by secondary medullary rays, which were either uniseriate or bi-seriate. Cambium was clearly visible.

> A leaf trace developed singly from the primary vascular strand. It divided into many mesarch strands upon entering into the cortex. At the place of its origin the leaf trace was C-shaped.
3. Reproductive Organs of Cycadeoidea:-
> The Bennettitalean reproductive organs are designated as “flowers “. The flower buds in the plants were present in the axil of leaf bases. As many on 500 flower buds were present on a single trunk in species such as Cycadeoidea dartonii (=Monanthesia dartonii).
> In several species of Cycadeoidea all the flower buds were present on a trunk at almost the same stage of development. Some palaeobotanists believe that such a plant might have flowered only once during its lifetime. Except a few species (e.g. C. wielendii) the flowers in Cycadeoidea were bisexual.
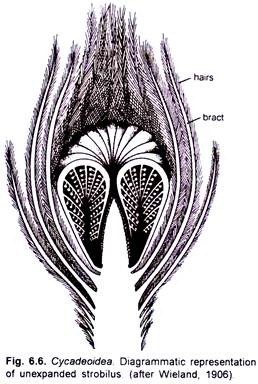
> Hermaphrodite flower developed on a short pedicel. They were surrounded by as many as one hundred bracts, which were hairy and protective. Flowers in different species were of different size. In Cycadeoidea dartonii they attained a length of about 2 cm and a diameter of about 1.5 cm while in C. dacotensis each flower was about 8 cm long and 3 cm in diameter.
> In C.dacotensis the lower two-third portion of the floral axis had about 100-150 bracts. A whorl of stamens was present above the bracts. Each stamen was pinnately branched and each pinna had a double row of purse-shaped sporangia. Each sporangium resembled with a synangium. A conical floral axis was present just above the whorl of stamens. The entire compact structure resembled with a strobilus.


4. Microsporophyll in Cycadeoidea:-
> According to Wieland (1906, 1916), the androecium or pollen-bearing region consisted of about 20 pinnate, microsporophyll’s. These were somewhat fixed or united at the base. Bean-shaped pollen capsules were arranged in two rows on each pinna of the sporophyll.
> These microsporophyll’s remained folded round the gynoecium when young, but probably at maturity they expanded. Delevoryas (1963), however, opined that the microsporophyll’s never expanded.
> He further concludes that synangia-bearing structures, described as pinnae by Wieland (1906), were similar to the trabeculae. These trabeculae established a connection between outer and inner walls of the androecium. Pollen capsules or synangia were borne along these trabeculae.
> Several (20-30) pollen sacs or microsporangia were present in a pollen capsule or synangium. The wall of a synangium consisted of outer palisade-like, thick-walled cells followed by thin-walled layer and then a tapetum.
> The tapetum was not clearly demarcated. The pollen grains were oval in shape and measured up to 68fi in length. Multicellular pollen grains in Cycadeoidea have been reported by Taylor (1973).


5. Gynoecium of Cycadeoidea:-
> The gynoecium receptacle was spherical or conical in shape. Hundreds of the stalked ovule along with an approximately equal number of inter-seminal scales were present on the receptacle (Fig. 6.10). Each ovules was about 1 mm in length. The integument of the ovule was fused with the nucellus, except at the apex.
> The ovule was orthotropous with a long micropylar beak. A pollen chamber and a nucellar beak was present in each ovule. The seed was somewhat elongated or oval in shape and possessed two cotyledons. Crepet and Delevoryas (1972) reported a linear tetrad in the nucellar region of Cycadeoidea.


